研究概要
研究テーマ:酵母と植物の Ca2+ 透過性伸展活性化チャネルの分子生物学的研究
キーワード:カルシウム、伸展活性化チャネル、機械受容チャネル、
Ca2+ チャネル、Ca2+ シグナル、シグナル伝達、 MAP キナーゼ、
カルシニューリン、細胞極性、酵母、シロイヌナズナ
研究の背景と目的(時間のない方はこの章だけお読み下さい)
動物も植物も接触や重力を敏感に感じることができます。それができる理由の一つは細胞膜に存在する Ca2+ 透過性の伸展活性化チャネル(stretch-activated channel; 略称 SAチャネル)または機械受容チャネル(mechanosensitive channel; 略称 MSチャネル)と呼ばれるイオンチャネルがあるからです。細胞膜がそれらの刺激によって伸びると(つまり伸展すると)、SA チャネルが膜の伸展を感知して開口し、細胞質に Ca2+ を流入させます(図1)。流入した Ca2+ は、膜の脱分極を引き起こすとともに Ca2+ シグナルとして酵素や遺伝子の活性化を引き起こします。その結果、細胞の興奮、増殖、分化、伸長などが起こります。動物の個体レベルでは、聴覚、平衡感覚、血圧など、植物の個体レベルでは、重力屈性やつるの巻付きなどが制御されます。
このように重要なはたらきをもつSAチャネルの構造と機能は長い間分子レベルでは分かっていませんでした。その理由はSAチャネルの遺伝子が不明だったからです。そのような状況の中で、我々は真核生物において世界で初めて酵母のSAチャネルを構成するタンパク質候補の遺伝子(MID1)を特定することに成功しました。また、その遺伝子に類似のシロイヌナズナ遺伝子を特定する努力を続けています。また、MID1 遺伝子産物(Mid1)と協力してはたらく Cch1 の研究も行っています。 Cch1 は動物の電位依存性Ca2+ チャネルの α1 サブユニットと構造上相同性があり、Mid1 との協調の観点からたいへん興味深い Ca2+ チャネル候補です。
以上の背景を基盤として、現在我々は以下の項目を明らかにすることを目的として研究しています。(1) Mid1の膜貫通領域、(2) Mid1の N-末端および C-末端領域の機能、(3) Mid1 と相互作用するタンパク質の特定と機能、(4) Cch1 の構造と機能、(5) Mid1 とCch1 の協調的機能、(6) Mid1 とCch1 による Ca2+ シグナルの発生機構と伝達機構、(7)シロイヌナズナの Mid1 と類似機能をもつタンパク質の特定と機能解析。これらの研究では分子生物学、細胞生物学、遺伝学、形態学などの方法が使われています。
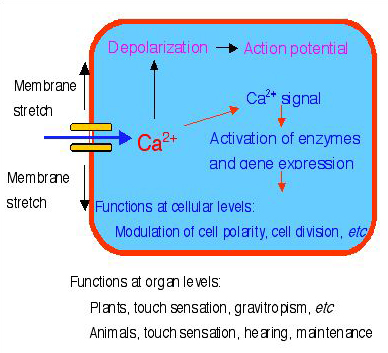
図1 伸展活性化Ca2+チャネルの役割
伸展活性化Ca2+チャネルが細胞膜の伸展により開口すると、細胞内にCa2+が流入する。
そのCa2+は細胞膜の脱分極とCa2+シグナルの発生を引き起す。
(興味を持った方は以下もお読み下さい)
酵母の特徴と有用性
基礎生物学で良く使われる酵母には、出芽酵母(Saccharomyces cerevisiase)と分裂酵母(Schizosaccharomyces pombe)があります。どちらもヒトも含めた真核生物の生命現象を分子レベルで解明するためのモデル生物として有用です。そのため、世界中で多くの研究者が酵母を使ってさまざまな生命現象を研究しています。私の研究室では、出芽酵母を使っています。
出芽酵母にはa細胞とα細胞と呼ばれる一倍体細胞があります。両者は性接合により
a/α 細胞(二倍体)になります。性接合は a 細胞と α 細胞がそれぞれ分泌する a 因子と α 因子と呼ばれる性フェロモンによって誘導されます。 a 細胞は a 因子の作用によって shmo oと呼ばれる接合突起を持った細胞に分化します(図2)。一方、α 細胞は a 因子の作用によって、shmoo に分化します。そして、それぞれの接合突起の先端で性接合を行い、二倍体細胞になります。性フェロモンの作用機構は、ほ乳類におけるホルモンの作用機構に似ていますので、盛んに研究が進められました。その結果、たとえば、G タンパク質やMAPキナーゼ経路のはたらきが明らかになっています。しかし、性フェロモンの作用後のCa2+ シグナルの発生のメカニズムは、たいへん興味深いのですが、まだ分らないことが多く残っています。我々の研究室はこの問題の突破口を切り開いてきた研究室の1つであり、幾つかの優れた実績と今後の発展性があります。
学生やポスドクにとって酵母を使うことの有意義な点の1つとして、酵母はモデル生物なので、酵母だけに留まらず、植物や動物の生命現象を同時に学びながら研究できることが挙げられます。
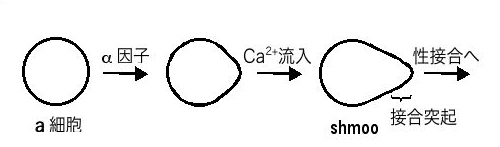
図2 出芽酵母におけるα因子によるa細胞の細胞分化
接合突起を形成しつつあるshmoo細胞の細胞膜では、膜の伸展によりMid1が活性化されている可能性がある。
Mid1 の構造と機能
Mid1は548アミノ酸残基からなり、N-グリコシレーション化され、細胞膜および小胞体膜に存在するタンパク質です(図3)。MID1 遺伝子を欠損した細胞(mid1Δ)は、(1) α 因子の作用を受けて shmoo に分化した後死ぬ、(2) Ca2+ 取込み活性が低い、(3) 静止期で死に易い。という表現型をもちます。また、ほ乳類細胞に発現させると、Mid1 は SA チャネル活性をもちます。この研究は我々の研究であり、真核生物の SA チャネルに関する分子レベルの研究としては世界で初めてのものです。
Mid1 は構造上幾つかの特徴を持っています。その代表的なものとして、第一に、4つの疎水性領域があります(図4)。N-末端から順にH1、H2、H3 および H4 と名付けられており、H1 領域は膜移行に必要なシグナル配列と予想されています。H2、H3 および H4 をそれぞれ単独で欠失させると Mid1 は完全に機能を失います。また、それぞれの領域内の幾つかのアミノ酸残基は Mid1 の機能に必須です。第二に、H4 領域下流の C-末端にはシステイン残基が多い領域があります。第三に、Mid1 は S-S 結合により複合体(おそらく二量体)を作っています。
今後の重要な課題として、Mid1の膜トポロジーの決定とC-末端領域システイン残基の役割の解明があります。
また、Mid1 により発生した Ca2+ シグナルの生理的役割を解明することも重要な課題です。この Ca2+ シグナルを受け取るのは、カルモデュリンとカルシニュリンであると予想されます。なぜなら、これらのタンパク質をコードする遺伝子を欠損した株は、mid1Δ株と類似の表現型の1つを示すからです。また、細胞壁合成に関与する MAP キナーゼ(MAPK)経路の活性化にもこの Ca2+ シグナルは関与している可能性があります。その理由は、同様にこの経路を構成する MAPK、MAPKK および MAPKKK の欠損株も mid1Δ株と類似の表現型の1つを示すからです。
Mid1 と直接的または間接的に相互作用して、Mid1 の機能を制御するタンパク質を同定することも重要な課題です。その幾つかを我々は明らかにしつつあります。たとえば、細胞極性の制御因子である Spa2 タンパク質はMid1の活性を正に制御していることを明らかにしました。その他の因子も研究中です。
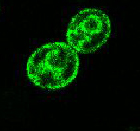
図3 Mid1タンパク質の局在部位
Mid1の局在部位を見るために、Mid1のC-末端に緑色蛍光タンパク質(GFP)を融合させ、
共焦点レーザー蛍光顕微鏡で観察した。Mid1-GFPは細胞膜と小胞体膜に局在することが明らかになった。
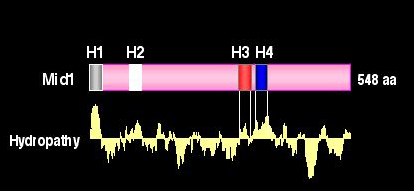
図4 Mid1タンパク質の構造
Mid1は548アミノ酸残基からなり、4つの疎水性領域(H1〜H4)をもつ。
Mid1 と Cch1 の協調
mid1Δ株、cch1Δ株および mid1Δ cch1Δ株の α 因子作用後の表現型は同じです。このことから、α 因子作用後において Mid1 と Cch1 は同じ Ca2+ チャネル系を構成している可能性が考えられます。一方アミオダロンという不整脈治療薬の作用後の Ca2+ 流入には、Cch1 と協調せず Mid1 が関与することが知られています。したがって、Mid1 とCch1 が細胞の置かれている状況に応じてダイナミックに協調関係を変えていることが予想されます。
Cch1 の遺伝子は大腸菌に毒性を示すので長い間クローニングをすることができないとされてきました。しかし、我々はプラスミドを工夫してこの遺伝子をクローニングすることに成功しました。現在、MID1 遺伝子とCCH1 遺伝子の両方を酵母細胞中に高発現させて、Mid1 とCch1 から成るチャネル系の構造と機能を調べています。先に述べましたように、Cch1 は動物の電位依存性 Ca2+ チャネルの α1 サブユニットと相同ですので、我々によるCch1 の分子生物学的研究は Ca2+ チャネルの研究分野全体の発展に大きく貢献をできるものと思います。
シロイヌナズナの Ca2+ 透過性 SA チャネル
植物において SA チャネル(または MS チャネル)は、機械刺激(たとえば接触刺激、重力刺激および浸透圧変化)のセンサーとしてはたらいていると予想されています。しかし、植物においてSAチャネル遺伝子とその産物であるチャネルタンパク質は未だ特定されていません。我々は酵母のMID1 と類似の機能をもつ候補遺伝子をシロイヌナズナ(図5)から単離し、解析しています。この研究は現在良い成果が挙げられつつあります。今後の詳しい解析により、機械刺激の受容の分子機構を解明できるものと期待されます。

図5 シロイヌナズナの野生株
シロイヌナズナ(Arabidopsis
thaliana)は、小さいサイズ(30 cm)、
短い世代時間(3ヶ月)、小さいゲノム(26,000遺伝子)という便利な特徴をもつ。
しかも、その遺伝子や変異体のコレクションが充実している。
そのため、シロイヌナズナはモデル植物として広く使われている。