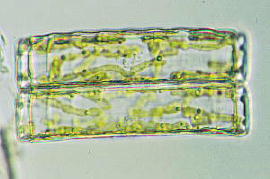
珪藻の葉緑体
![]() 光合成色素
光合成色素
珪藻の葉緑体はクロロフィルaとc1、c2を含んでいます。そして,緑色植物(クロロフィルaとbを含む緑藻~車軸藻~コケ~シダ~種子植物という系統の生物)とは全く別系統の黄色植物に属します。光学顕微鏡で観察すると黄色に見えるのは葉緑体にクロロフィルのほかに、ディアトキサンチン、ディアディノキサンチン、フコキサンチンといった黄色い色素を含むためでです。緑色に見えないため色素体という言葉を好んで使う研究者たちも多くいます。珪藻の黄色い色素は熱、酸、アルコール等に弱くそれらの影響を受けると、葉緑体は緑色になりますので,やはり珪藻もクロロフィルaを含む藻類であることがわかります。
![]() 数と形
数と形
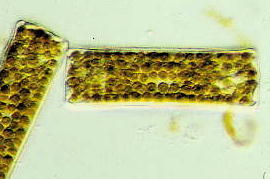
葉緑体の数は中心珪藻では1細胞中に多数もつ種類が多く、羽状珪藻の縦溝類では1細胞中に1, 2枚持つものが多い傾向にあります。羽状珪藻の無縦溝類はこれらの中間で,多数もつ種類と少数持つ種類が同じような割合でみられます。葉緑体の形は多様です。最も単純な形は板状ですが,このほか多角形、蝶形、紐形、不定形、楕円形、円盤形、顆粒状のものながあります。しかし葉緑体について集中的に行った研究は非常に少なく、過去における総説はなんと20世紀初頭の1902-1903年にメレシュコフスキー (C.Mereschkowsky) が執筆したものと、1908年にハインツァーリンク (O.Heinzerling) によるものしかないのです。
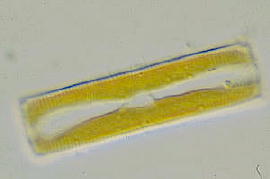
![]() 分裂様式
分裂様式
葉緑体の分裂は、核分裂に先立って行われるものと、核分裂後娘細胞が形成されてから分裂するものとがあります。葉緑体分裂の様式は縦に割れるもの、横に割れるもの、斜めに割れるものなどがあり、また分裂に際し葉緑体の移動や回転がおきることもあります。しかし,分裂様式の報告例は決して多くはありません。

![]() 分類への応用
分類への応用
珪藻の分類に葉緑体を利用しようとしたのはメレシュコフスキーでした。彼は,葉緑体の形状に基づき新属を設立したり分類の体系化を試みています。しかし,彼の意見は重要視されず,その後80年以上も珪藻の分類は殻の構造のみによって行われ続けました。
ところが最近になり,葉緑体の形状が再び注目されてきました。それは、葉緑体の形状や分裂様式が同じ種類において殻を観察すると,その微細構造にも共通の形態があることがわかってきたためです。たとえば、フネケイソウ属 (Navicula)に分類されていたいくつかの種類はセラフォラ属 (Sellaphora)に移されました。Navicula属の種には板状の葉緑体が2枚ありますが,Sellaphora属の種類にははH形の葉緑体が1枚しかありません。また,Sellaphora属の種類には殻の両極に洞穴状の独特の構造(cavity)があるのですが,Naviculaにはこのような構造はありません。
![]() 葉緑体微細構造とピレノイド
葉緑体微細構造とピレノイド
珪藻の葉緑体は3重チラコイドラメラからなり、葉緑体周辺を囲むガードルラメラが存在します。葉緑体中にピレノイドはおそらくほとんどの種類に含まれていると思われますが、その数は1個の場合もあれば、数個の種類もあるようです。レンズ形をしてチラコイドが1枚貫通しているピレノイドは珪藻における典型的なものです。しかし,それ以外の形をとる種類の存在も知られています。ピレノイドの数や位置は他の藻類では重要な分類形質になっている場合がありますが、珪藻ではまだ限られた種類についてのみ観察されているのが現状で、ピレノイドの研究は今後の課題の一つとなっています。
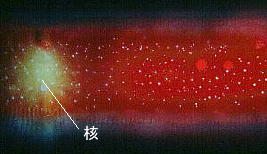
![]() 葉緑体核様体
葉緑体核様体
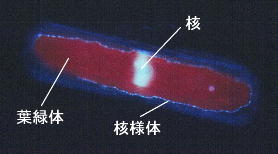
近年全ての植物の葉緑体にはDNAが含まれることが知られるようになりました、珪藻の葉緑体DNAの分布は知られている限りでは葉緑体周囲にリング状に配置されています。これはDNAを特異的に染色するDAPI (4'6-diamidino-2phenylindole)試薬と落斜型蛍光顕微鏡を用いることで観察することができます。しかし、まだ観察報告されている珪藻種はほんのわずかです。私たちはPinnularia nobilisには葉緑体周囲に分布するDNAの他に,葉緑体全域にわたり分散するDNA含有顆粒の存在を明らかにしましたが,これも核様体分布の一様式であると考えています。
![]() 葉緑体の光・接触刺激に対する挙動
葉緑体の光・接触刺激に対する挙動
葉緑体を多数持つ種類では、昼と夜で葉緑体が細胞内を移動する種類もあります。これらの種類では昼間葉緑体は被殻の真下に広がっているのですが,夜間は核の周辺部に集まります。また,実験的に強い光を当てても葉緑体は夜間同様に核周辺部に集まります。さらに,当てる光の色を変えると異なる動きが観察されます。葉緑体の細胞内移動は接触刺激でも起こることが近年Pleurosira laevisでわかりました。また,この刺激は離れた個体にも伝搬することも実験的に確かめられています。
![]() 葉緑体ER
葉緑体ER
緑色植物の葉緑体は2枚の膜によって包まれています。ところが珪藻の葉緑体は4枚の膜によって包まれているのです。これは、珪藻を含む黄色植物すべてに共通する特徴です。この4枚の膜のうち、外側の2枚を葉緑体ER (chloroplast endoplasmic reticulum) と呼び、内側2枚を葉緑体膜 (chloroplast envelope) と呼びます。珪藻では葉緑体ERの外膜は,核膜の外側の膜とつながっています。また,葉緑体膜の外膜と葉緑体ERの内膜の間に,葉緑体周縁区画(ペリプラスチダルスコンパートメント)と呼ばれるわずかな間隙が認められます。
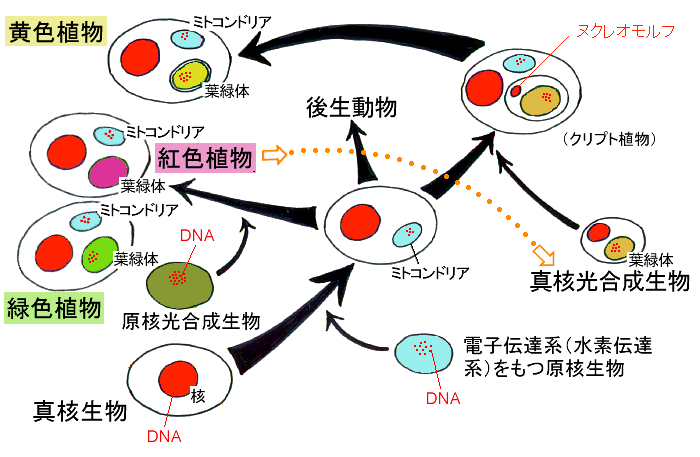
![]() 二次共生による葉緑体の獲得
二次共生による葉緑体の獲得
珪藻を含む黄色植物の葉緑体は宿主細胞内に共生した「真核」光合成生物が,その起源と考えられています。真核光合成生物はすでに「原核」光合成生物を起源とする葉緑体を共生の結果獲得していますので,黄色植物は二次共生の結果葉緑体を獲得した生物といえます。ここで葉緑体の膜と共生進化について考えてみましょう。
←クリックすると拡大されます
4枚あるうち,もっとも内側の2枚は共生体(真核光合成生物)の葉緑体膜です。ここで,問題となるのが2枚の葉緑体ERです。まず,葉緑体ERの内膜ですが,これは共生体の細胞膜であったと考えられます。そして葉緑体ERの外膜は共生体を取り込んだ宿主の食胞膜と考えられます。そして,葉緑体周縁区画はその昔,共生体の核や細胞質の詰まっていた場所と考えられます。また,葉緑体膜の外膜と葉緑体ERの内膜の間にある葉緑体周縁区画は,かつて共生体の核や葉緑体以外の細胞質が存在した場所の痕跡と考えられているのです。4枚ある葉緑体の膜には,葉緑体獲得にまつわる共生進化の歴史が秘められているのです。
「珪藻の世界」ホームページへ